link to page 12 link to page 14 link to page 14 link to page 12 link to page 13 link to page 13 link to page 14 link to page 14 link to page 12 link to page 12 link to page 12 link to page 13 link to page 14
ForestryAnInternationalJournalofForestResearch
Forestry 2019; 92, 1–15, doi:10.1093/forestry/cpy024
Advance Access publication 3 August 2018
Assessment of multiple climate change effects on plantation forests
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
in New Zealand
Michael S. Watt1*, Miko U. F. Kirschbaum2, John R. Moore3, H. Grant Pearce1, Lindsay S. Bulman3,
Eckehard G. Brockerhoff1 and Nathanael Melia3
1Scion, PO Box 29237, Christchurch 8011, New Zealand
2Landcare Research, Private Bag 11052, Palmerston North 4442, New Zealand
3Scion, Private Bag 3020, Rotorua 3046, New Zealand
*Corresponding author Tel: +64 3 363 0910; E-mail: [email address]
Received 21 March 2018
Determining the magnitude of climate change effects is crucial for informing national economic strategies,
forest management and offsetting increasing carbon emissions. This study synthesizes predicted climate
change impacts and future biosecurity threats to New Zealand’s plantation forests. Projected productivity
increases for radiata pine (Pinus radiata D. Don), the main commercial forest species in New Zealand, are slight
due to changing climatic conditions. However including photosynthetic effects from increasing CO2, productiv-
ity gains across New Zealand averaged 19 per cent by 2040 and 37 per cent by 2090. This increased product-
ivity results in marked increases in wind risk due to trees becoming taller and more slender. The average
season length with ‘very high and extreme’ climatic fire risk increases by 71 per cent up to 2040 and by 83
per cent up to 2090. Currently, the most significant biotic disturbances in New Zealand plantations come from
two needle cast diseases, for which climate projections show slight increases or decreases depending on the
disease and region. Although insect pests currently cause little damage to New Zealand plantations, damage
may increase in the future with projected increases in population and host susceptibility. It has not been pos-
sible to fully account for the effects of any new introductions of pests and pathogens and evidence from other
countries with a significant resource of planted forests suggest this should not be underestimated. Potentially
invasive weedy and damaging tree species are likely to expand their range under climate change and com-
pete more strongly with plantations.
Introduction
around the magnitude of this response. Photosynthesis of C3
plants, including trees, increases strongly up to an ambient CO2
Climate change is emerging as one of the key influences to
concentration of about 300–400 ppm. Current observations show
shape the future of natural and anthropogenic systems across
CO2 rising above 400 ppm
(Betts et al., 2016), and with increasing
the world
(Cramer et al., 2014). As forests cover about one quar-
CO2 concentrations, relative photosynthetic gains by ongoing
ter of the Earth’s land surface area, they play a major role in
increases in CO2 concentrations become progressively smaller
current and projected future carbon budgets. Determining the
(
Kirschbaum, 2011; Hickler et al., 2015).
influence of climate change on planted forests is of importance
However, while there may be gains in productivity realized
as these forests provide about half of the global wood supply
through increased CO2 and temperature up to the optimal value,
and are a vital natural mechanism to offset future carbon emis-
they may be counteracted to varying degrees by changes in abi-
sions (
Payn et al., 2015). Increased disturbances to forests may
otic and biotic stressors
(Reyer et al., 2017). Climate change will
reduce carbon stocks, result in substantial economic impacts
influence the distribution and abundance of many forest pests
and have consequences for ecosystem functioning.
(Unless otherwise stated (i.e. insect pest), pest refers to all agents
Considerable research has investigated how climate change
injurious to trees or tree products (i.e. insects, pathogens and
influences forest productivity. A recent review that synthesized
weeds)) and alter the frequency and intensity of damaging abiotic
findings from 31 studies found that 87 per cent of them identi- factors, such as wind and fire (e.g.
Walther et al., 2009; Allen et al.,
fied positive changes in forest productivity when the effects of
2010; Anderegg et al., 2015). Predicted changes in these factors
climate change and increasing CO2 were combined (
Reyer, 2015).
under climate change have often been found to lead to reduced
Although gains in forest productivity are likely to occur as a result
productivity
(Kurz et al., 2008; Seidl et al., 2014). Although a com-
of climate change and CO2 increases, there is some uncertainty
prehensive assessment of changes in productivity has to account
© Institute of Chartered Foresters, 2018. All rights reserved. For Permissions, please e-mail: [email address].
1
link to page 12 link to page 15 link to page 14 link to page 14 link to page 13 link to page 13 link to page 13 link to page 15 link to page 15 link to page 15 link to page 15 link to page 13 link to page 13 link to page 3 link to page 3 link to page 3 link to page 14 link to page 13 link to page 14 link to page 14 link to page 14 link to page 13 link to page 14 link to page 13 link to page 13 link to page 14 link to page 13 link to page 13 link to page 13 link to page 13 link to page 13 link to page 13 link to page 13 link to page 13
Forestry
for the impacts of changes in all these factors, combined assess-
the far north and east coast of the North Island, the upper South
ments such as in this paper have rarely been done in the past (e.g.
Island and various locations along the east coast of the South
Chen et al., 2000;
Wolken et al., 2011; Shanley et al., 2015).
Island, especially in the far south (Figure
1c).
New Zealand provides a useful case study for examining the
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
overall effects of climate change on plantation productivity as a
Materials and methods
single species, Pinus radiata D. Don, covers 90 per cent of the 1.7
million hectare plantation resource
(NZFOA, 2016). There is much
Climate change projections
information describing the physiology and morphology of P. radia-
Values of meteorological variables were estimated for the whole of New
ta, and how this species responds to changes in environmental
Zealand on a 0.05° latitude/longitude (≈5 × 5 km) grid, using a thin-
factors. Using this information, previous research has modelled
plate smoothing spline to spatially interpolate daily observational data
the growth of P. radiata under current (
Kirschbaum and Watt,
(Tait et al., 2006;
Ministry for the Environment, 2008;
Tait, 2008; Tait and
2011) and future
(Kirschbaum et al., 2012) climatic conditions
Liley, 2009). Climate change projections used in this study were derived
within New Zealand. The major needle diseases of P. radiata are
from the factorial combination of 12 Global Climate Models (GCMs) and
well known, and the spatial variation in the severity of two prob-
the SRES emission scenarios, B1 (low), A1B (mid-range) and A2 (high),
lematic diseases (caused by Dothistroma septosporum (Dorogin)
described by
Meehl et al. (2007). The B1, A1B and A2 scenarios approxi-
M. Morelet and Cyclaneusma minus (Butin) DiCosmo, Peredo and
mately correspond to the newer representative concentration pathways
Minter) has been described under both current and future cli-
4.5, 6.0 and 8.5 (
Rogelj et al., 2012). The 12 GCMs used in this study had
matic conditions (
Watt et al., 2011a, b, 2012a, b). Spatial vari-
been selected because of their utility in modelling 1971–2000 climatic
conditions (mean sea level pressure, temperature and precipitation) over
ation in wind risk under current and future climate has also been
New Zealand from the widely used NCEP reanalysis
(Kalnay et al., 1996;
quantified
(Moore and Watt, 2015).
Ministry for the Environment, 2008). They were as follows: CNRM,
The primary aim of this study was to synthesize knowledge
CCCma, CSIRO Mk3, GFDL CM 2.0, GFDL CM 2.1, MIROC32, ECHOG,
describing the likely impacts of climate change on P. radiata plan-
ECHAM5, MRI, NCAR, UKMO-HadCM3 and UKMO-HadGEM1. Monthly tem-
tations. Specifically, we summarize previously published studies
perature and rainfall scenarios from each GCM were statistically down-
describing the impact of climate change on radiata pine product-
scaled to a resolution of 0.05° for 1990 (henceforth ‘baseline’), 2040
ivity and damage from wind and two of the major needle dis-
and 2090, using the methods of
Mullan et al. (2002).
eases of P. radiata. We complement this synthesis with additional,
previously unpublished, research describing the potential impacts
Modelling forest productivity
of fire on plantations. A general more qualitative synthesis of the
literature was undertaken to describe the potential impacts of
The process-based model CenW version 4.0 (
Kirschbaum, 1999) was used
to project productivity of P. radiata across New Zealand under current and
biotic factors that includes a key report describing changes in
future climates (
Kirschbaum and Watt, 2011; Kirschbaum et al., 2012).
trade patterns and their effects on the origin of biosecurity risks
CenW has been developed primarily for climate change investigations
(Kean et al., 2015). We describe the potential future impacts of
and incorporates the key processes and feedbacks between plants and
key diseases, weeds and insects that are either currently in the
their environment that can operate on timescales ranging from daily (for
country or could pose major incursion risks. We conclude with
water relations) to decadal and longer (for soil organic matter feedbacks
sections describing key sensitivities highlighted by the study and
and wood growth).
Kirschbaum and Watt (2011) demonstrated that
areas for further research.
CenW can successfully model stand productivity of P. radiata under cur-
rent climatic conditions within New Zealand, providing confidence that
the model incorporates the key processes underpinning productivity.
New Zealand’s climate and the location
CenW was parameterized for current climatic conditions using
growth data from permanent sample plots covering almost the com-
of plantations
plete environmental range across which P. radiata is grown in New
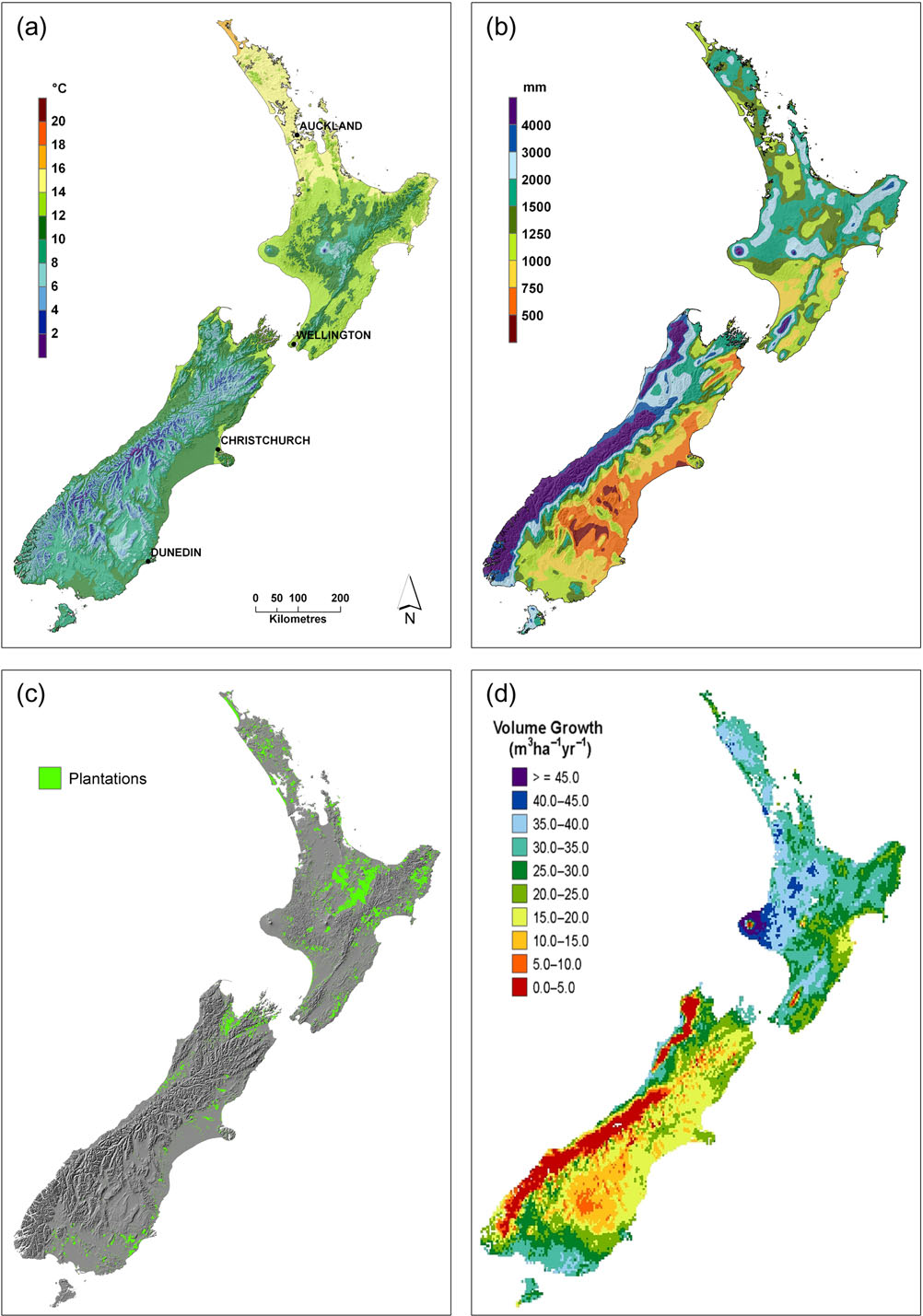
New Zealand’s mean annual temperature at low-elevation
Zealand. These data consisted of 101 sites with 1297 individual observa-
tions of height and/or basal area from which diameters and volumes
sites ranges from 8°C in the south to 16°C in the north, with
were calculated. Using the parameter values determined by
Kirschbaum
colder conditions at higher elevations, especially in the South
and Watt (2011), there was excellent correspondence between model
Island (Figure
1a). Variation between summer and winter tem-
predictions and measurements of a range of tree dimensions.
peratures is generally relatively small, especially in coastal
Simulations were run under both current and future climatic condi-
regions. Consequently, there are currently few periods with
tions using projections from the 12 GCMs and the three emission scen-
extremely hot or cold conditions in the low to moderate eleva-
arios described previously. Future climate scenario outputs were used in
tion areas of New Zealand where plantation forests are grown.
CenW with both constant and increasing CO2 to isolate tree responses
Precipitation within most of New Zealand ranges from 500–2000
to changing climatic conditions and elevated CO2. The results presented
mm yr −1 (Figure
1b). Mountain ranges extending throughout New
hereafter are the mean of the 12 GCMs unless otherwise stated.
Zealand provide a barrier to the prevailing westerly winds, dividing
the country into markedly different climatic regions. The west coast
Wind damage
of the South Island is the wettest area of New Zealand, with a num-
The risk of wind damage was quantified using the approach described
ber of locations receiving over 5000 mm yr−1, whereas the area to
by
Moore and Watt (2015). They investigated both the direct effects of
the east of the mountains, just over 100 km away, is the driest,
increasing wind speeds and the indirect effects of changes in stand
with annual precipitation reaching minima of 500 mm yr−1.
structure, which affects the underlying susceptibility to wind damage.
New Zealand’s exotic plantation estate is distributed through-
These impacts were investigated using representative growth rates and
out most of the country, with the largest areas in the central
climatic conditions for seven bio-geo-climatic zones for P. radiata in New
North Island. Substantial areas of plantation are also found in
Zealand
(Goulding, 1994). Site productivity metrics for these zones were
2
link to page 13 link to page 14 link to page 15 link to page 13
Climate change effects on plantation forests
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
Figure 1 New Zealand maps of current (1980-1999) (a) mean annual temperature and (b) mean annual rainfall (after
Wratt et al. (2006)), (c) the
current location of plantation forests and (d) modelled wood productivity (as volume growth) of P. radiata under current climatic conditions (redrawn
from
Kirschbaum & Watt, 2011).
used to predict the stand structure (diameter, height, volume and spa-
speed required to damage mean trees within a stand. The average
cing) for three contrasting silvicultural regimes (pruned, unpruned and
annual probability that these critical wind speeds were exceeded was
carbon) under current and future climatic conditions. Stands grown
estimated from frequency distributions of extreme wind speeds calcu-
under a carbon regime are planted at a very high stand density and left
lated from time series of observations from long-term meteorological
in perpetuity with the sole purpose of maximizing carbon sequestration.
stations in each zone.
This information was then input into a mechanistic wind damage model,
Although there is still considerable uncertainty around New Zealand’s
ForestGALES (
Gardiner et al., 2000), in order to predict the critical wind
future extreme wind climate, an analysis carried out by
Mullan et al. (2011)
3
link to page 14 link to page 14 link to page 12 link to page 14 link to page 14 link to page 13 link to page 14 link to page 14 link to page 14 link to page 14 link to page 14 link to page 15 link to page 15 link to page 15 link to page 15 link to page 8 link to page 15 link to page 15 link to page 4 link to page 4 link to page 5 link to page 5 link to page 3
Forestry
has indicated that extreme wind speeds are only likely to increase by
Table 1 Summary of the mean simulated changes in temperature,
between 1 and 5 per cent under the A1B scenario, with no predictions of
precipitation and CO2. GCM minimum (min) and maximum (max) refer
any changes available under other future scenarios. We accounted for these
to the GCM with the lowest and highest spatially averaged changes.
potential increases through increasing the mode of the extreme wind speed
Variation of simulated changes between the GCMs is also expressed as a
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
distribution by 5 per cent for all simulated time periods
(Quine and Gardiner,
standard deviation (SD) which expresses the variation between the
2002). All results are expressed at the age of 30 years as an annual excee-
average NZ values between the 12 GCMs.
dance probability (AEP) which is defined as the likelihood of a damaging
wind event occurring in a given year, with values ranging from 0 to 1.
Year
Emission CO2
Temperature
Rainfall
scenario
(ppm) change (°C)
change (%)
Fire risk
Fire danger ratings (using 1970–1999 as a baseline) were computed using
Mean GCM
Mean GCM
temperature, humidity, wind speed and rainfall data from the A1B emis-
Min Max SD
Min
Max SD
sions scenario, previously described. These changes were applied to 20
weather station sites across New Zealand with at least 20 years of obser-
2040 B1
457
0.7
0.4
1.2
0.3 1.6
–2.1
5.6 2.3
vations to calculate future daily Fire Weather Index (FWI) and fire danger
A1B
483
0.9
0.4
1.3
0.3 2.1
0.0
5.0 1.6
class values
(Anderson, 2005). Fire risk is classified as being Low,
Moderate, High, Very High, or Extreme. The fire climate severity is quanti-
A2
481
0.9
0.3
1.3
0.3 0.9
–5.6
4.3 2.6
fied here as the frequency of days in each fire season that had Very High
2090 B1
538
1.3
0.7
2.3
0.4 2.6
–4.7
7.9 3.8
or Extreme (VH + E) fire danger. Further details of the methods used in
A1B
674
2.1
1.2
3.4
0.6 3.2
–1.5 12.9 3.9
this study are described by
Pearce et al. (2011) and
Simpson et al. (2014).
A2
754
2.6
1.6
3.6
0.5 3.0
–3.8 11.5 5.6
Projected changes in fire danger for the whole of New Zealand were
produced by spatially interpolating the changes predicted at each of the
20 station locations using the co-kriging technique
(Goovaerts, 1999). The
co-kriging technique allowed inclusion of additional surface prediction
scenarios by 0.7−0.9°C by 2040 and by 1.3−2.6°C by 2090,
variables for station location (latitude/longitude), elevation and informa-
although the inter-model uncertainty outweighs the scenario
tion from additional stations in data sparse locations
(Pearce et al., 2011).
uncertainty (Table
1). Projected multi-model mean increases in
rainfall for New Zealand under the three emission scenarios were
Disease damage
between 0.9 per cent − 2.1 per cent by 2040 and 2.6 per cent
−3.2 per cent by 2090, with significant inter-model variation with
Damage from foliar pathogens is currently the most costly natural disturb-
even the sign of the change differing between GCMs implying con-
ance to New Zealand plantation forests
(Watt et al., 2008). Dothistroma sep-
siderable uncertainty in regional and national precipitation projec-
tosporum, which causes dothistroma needle blight is currently the most
tions (Table
1). However, a consistent pattern in these simulations
damaging forest pathogen of P. radiata plantations. Phytophthora pluvialis
(Reeser et al., 2013), which is associated with red needle cast, was detected
was the absence of any extreme changes, with all model projec-
in 2008 and has the potential to cause significant damage within planta-
tions falling within the range between −5.6 and +12.9 per cent.
tions (
Scott and Williams, 2014). Cyclaneusma minus which results in cycla-
Changes under the A1B scenario showed the largest increases in
neusma needle cast is also important but of lesser concern. Other pine
rainfall by 2090, with changes under the A2 scenario falling
pathogens currently within New Zealand may cause sporadic or localized
between those under the B1 and A1B scenarios.
damage but their national impact is not significant
(Watt et al., 2008).
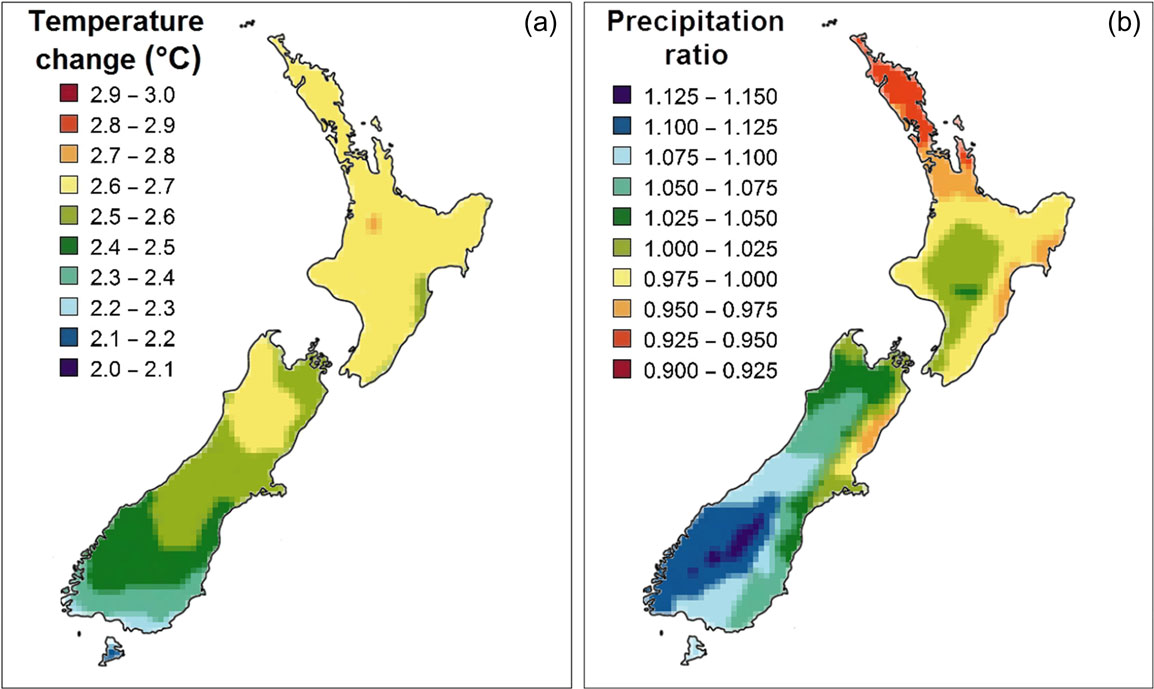
The mean 2090 projections for the A2 scenario (Figure
2) illus-
The approach used to determine damage from D. septosporum and C.
trate the regional patterns of the expected changes in tempera-
minus has been described in detail previously
(Watt et al., 2011a, b, 2012a,
ture and precipitation. Expected temperature changes broadly
b) and is briefly summarized in the following. Disease incidence and severity
correlated with latitude, with expected warming ranging from
data were collected from plantations throughout New Zealand over a 45-
2.7°C in the north to 2.2°C in the far south. Variation in precipita-
year period for dothistroma needle blight and over a 34-year period for
tion also showed a latitudinal correlation, with rainfall increasing
cyclaneusma needle cast. Disease severity, Ssev, was determined at the stand
in the already wet south-western and elevated areas, while rela-
level by multiplying the percentage of trees in the stand affected by mean
tively drier areas along the east coast and in the north are pro-
severity (percentage of needles affected) on affected trees (scale = 0 – 100).
Growth reduction generally occurs when disease severity exceeds 20 per
jected to become drier (Figure
2b).
cent, whereas disease severity values of 2–10 per cent (Table
3) are not likely
to significantly impair growth. However, stands and individual trees growing
in disease-prone regions may have values at which significant growth loss
Modelling forest productivity
occurs. Multiple regression models of Ssev were developed for both diseases
from meteorological data, described above, using the methods described
Under baseline climatic conditions, there was wide regional vari-
fully by
Watt et al. (2011b,
2012b). Using these multiple regression models,
ation in predicted stem volume growth (Figure
1d). Values were
spatial predictions of Ssev were made under current climate and to 2040 and
highest in the warm and moderately wet northern and western
2090 using the climate change scenarios previously described.
areas of the North Island, reaching maximum growth rates of
>40 m3 ha−1 yr−1 in the fertile Taranaki region. Productivity was
considerably lower within the South Island, partly attributable to
Impacts
cooler temperatures that were generally sub-optimal. Reduced
productivity was also attributable to excessively high rainfall
Climate change projections
(>3000 mm yr−1) on the west of the main axial mountain ranges
Averaged across the 12 GCMs, mean air temperature in New
and relatively low rainfall (<750 mm yr−1) on the eastern side. In
Zealand was projected to increase between the three emission
contrast, in the most southerly regions, where there is moderate
4
link to page 3 link to page 3 link to page 5 link to page 13 link to page 13 link to page 5 link to page 5 link to page 13 link to page 6 link to page 6
Climate change effects on plantation forests
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
Figure 2 Projected changes in temperature (a) and precipitation (b) for 2090 from baseline under the A2 emission scenario.
rainfall (Figure
1b) and little seasonal water deficit, productivity
was predicted to be higher than in regions with greater rainfall
extremes (Figure
1d) but still considerably lower than in the North
Island, owing to the much lower temperatures.
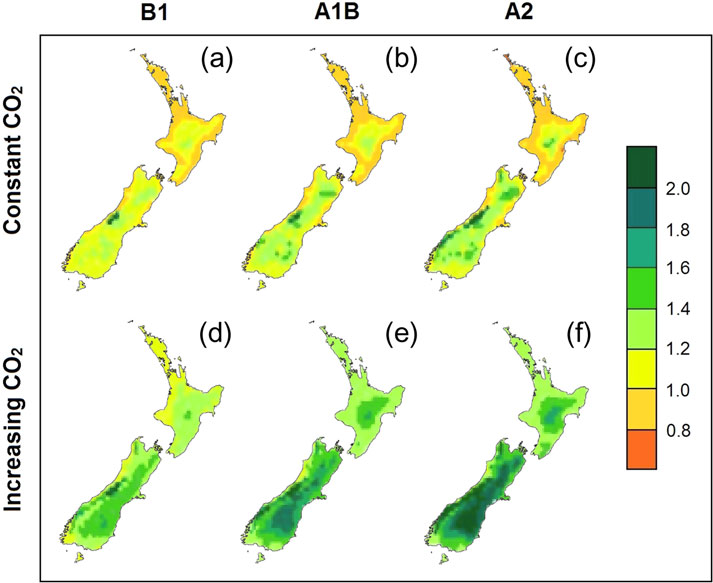
Neglecting future increases in CO2 isolates the effect of expected
climatic changes on future volume productivity. Projections to 2090
showed reduced productivity in northern and low-elevation regions
and increased productivity in southern and upland regions
(Figure
3a–c), likely due to shifts in temperature towards the opti-
mum range for P. radiata, which
Kirschbaum and Watt (2011) found
to be at a mean annual air temperature of 12–15°C. These changes
in climatic conditions resulted in growth gains for about half of all
plantations
(Kirschbaum et al., 2012), with mean changes in volume
productivity within plantation forests averaging +3 per cent by 2090.
Simulations with increasing CO2 led to productivity increases
during 2090 for all regions and under all emission scenarios
(Figure
3d–f). Increasing CO2 completely reversed the losses in
low-lying and northerly regions that had been modelled due to
climatic shifts. Productivity gains were predicted throughout
New Zealand and reached increases exceeding 100 per cent
within parts of the South Island (Figure
3d–f). Within the current
Figure 3 Mean volume productivity ratio in 2090 compared to current
plantation estate, simulations showed mean productivity
productivity with constant (top row) and increasing CO2 (bottom row)
increases to 2040 and 2090 of 19 per cent and 37 per cent,
under the B1 (a, d), A1B (b, e) and A2 (c, f) emission scenarios.
respectively (
Kirschbaum et al., 2012).
For a 30-year-old stand, i.e. at typical harvest age, the risk of
Abiotic impacts
wind damage was relatively low under the baseline climate.
Under this baseline, AEP ranged from 0.094 for the unpruned
Wind damage
regime to 0.166 for the carbon regime (Table
2). Projections to
The AEP, which is defined as the likelihood of a damaging wind
2040 for the pruned regime show that AEP was less than 0.2 for
event occurring in a given year, at the age of 30 years is given
all scenarios apart from the A2 emission scenario with CO2 held
for each silvicultural regime and emission scenario in Table
2.
constant at 1990 levels where the AEP was 0.286. Projections to
5
link to page 6 link to page 6 link to page 6 link to page 13 link to page 12 link to page 13 link to page 13 link to page 13 link to page 6 link to page 6 link to page 6 link to page 6 link to page 6 link to page 6 link to page 7 link to page 7 link to page 7 link to page 6
Forestry
2090 showed that the AEP increased on average by 0.066, and
AEP (Figure
4). Mean AEP ranged from 0.21 under the pruned
for four of the six scenarios, the AEP was >0.2 (Table
2).
regime to 0.24 under the unpruned regime and 0.50 under the
The risk of wind damage for the unpruned regime was very
carbon regime (Table
2). The growth response to increasing CO2
similar to the risk for pruned stands in both 1990 and 2040.
had relatively little impact on AEP in 2040 but a greater impact
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
However, for projections to 2090, the risk of wind damage for the
in 2090 at which time it was equal in importance to the silvicul-
unpruned regime was substantially higher than for the pruned
tural regime. Relative to other factors, both emissions uncer-
regime due to the higher ratio of height to diameter for this
tainty and increasing wind speed had very little effect on AEP,
regime. These increases were particularly marked for emission
and together, they accounted for less than 4 per cent of the
scenarios that assumed a full response to increasing CO2
total variance during both 2040 and 2090 (Figure
4).
(Table
2). The risk of wind damage was markedly higher for the
carbon regime than for the other two regimes due to the higher
height to diameter ratio of trees. The carbon regime had very
high sensitivity to increasing CO
Fire risk
2. For projections made to 2090,
scenarios that assumed a full response to increasing CO2 had an
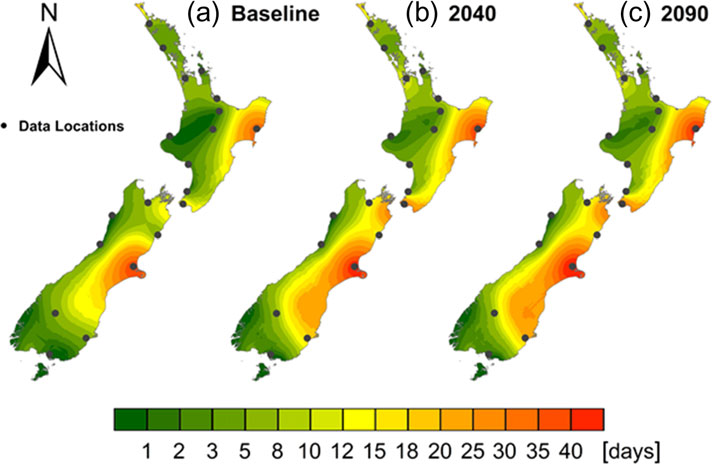
Under the baseline climate, dryland areas on the east coast had
AEP that was on average 0.33 higher than those that assumed
the highest average number of VH + E fire danger days per year,
no response, and the AEP of these three scenarios ranged from
while many areas on the west coast had a very few, or no VH + E
0.639 for the B1 to 0.922 for the A2 emission scenario (Table
2).
days. There was widespread spatial variation in the degree of
Stands grown on a carbon regime tended to be overstocked, par-
change in frequency of VH + E days between baseline and future
ticularly in the latter part of the rotation and the increased risk of
climate (Figure
5). The regions with the most notable VH + E
wind damage in such situations is consistent with experiences in
increases were located on the eastern coast in the southern half
other regions of the world (
Mitchell, 2000; Cameron, 2002).
of both islands.
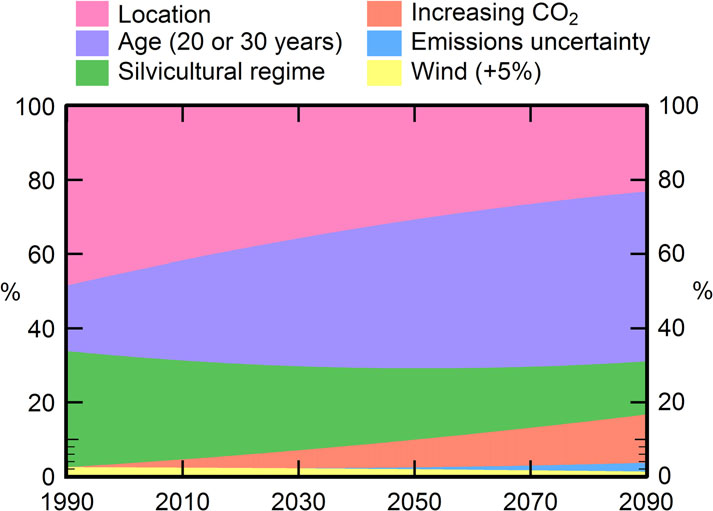
The relative contributions of different factors on AEP at 2040
Examination of the fire risk by location demonstrates the
and 2090 were determined using previously described methods
high variation between sites with the frequency of VH + E fire
(Hawkins and Sutton, 2009,
2011;
Melia et al., 2015). Figure
4
risk under the baseline climate ranging from 0 to 40 days
shows that currently, most of the variation in AEP is attributable
(Figure
6). When averaged over all sites, the number of days
to location, with stand age and silvicultural regime also being
with VH + E fire risk was projected to increase by 71 per cent by
important. Under future climates, mean values of AEP for a 30-
2040, and by a further 12 per cent by 2090. All sites on the east
year-old stand ranged from 0.18 to 0.79 across locations (data
coast showed increases under climate change. The locations
not shown). Stand age accounted for 27 per cent of the vari-
with highest current fire risk, Christchurch and Gisborne, had sig-
ance in AEP under future climates (Figure
4). The change in
nificant further increases in VH + E fire risk by 2090 to 44 and
mean AEP for increasing stand age from 20 to 30 years ranged
48 days, respectively. However, the most marked relative
from 0.09 to 0.33 in 2040 and from 0.12 to 0.49 in 2090.
changes occurred in Wellington (lower North Island) and
Silvicultural regime was relatively important (Figure
4) under
Dunedin (south-eastern South Island) where VH + E fire risk
future climates, accounting for 19 per cent of the variance in
increased to 2090 by, respectively, 89 per cent to 32 days and
207 per cent to 18 days (Figure.
6).
Table 2 AEP of wind damage in 30-year-old stands as a function of
simulation year, emission scenario and CO2 concentration for three
silvicultural regimes. Values shown represent the means across seven
bio-geo-climatic zones defined by their current wind climate. Values of
AEP are differentiated by colour into the categories of AEP < 0.20
(green), 0.2–0.5 (orange) and >0.5 (red).
Year
Emission
Inc.
Silvicultural regime
scenario
CO2
Pruned
Unpruned
Carbon
1990
0.110
0.094
0.166
2040
B1
N
0.152
0.141
0.262
2040
A1B
N
0.164
0.155
0.291
2040
A2
N
0.286
0.154
0.286
2040
B1
Y
0.150
0.172
0.419
2040
A1B
Y
0.164
0.201
0.507
2040
A2
Y
0.164
0.197
0.495
2090
B1
N
0.186
0.182
0.344
2090
A1B
N
0.238
0.242
0.443
2090
A2
N
0.272
0.278
0.483
Figure 4 Relative contribution of location, stand age, silvicultural regime,
2090
B1
Y
0.191
0.256
0.639
increasing CO2, emissions scenario and wind speed to AEP. Values of rela-
2090
A1B
Y
0.267
0.423
0.850
tive importance used in the figure were extracted from Table
2 and rela-
2090
A2
Y
0.321
0.522
0.922
tive importance was interpolated between years using second-degree
polynomials.
6
link to page 13 link to page 13 link to page 13 link to page 14 link to page 14 link to page 14 link to page 14 link to page 8 link to page 12 link to page 12 link to page 7
Climate change effects on plantation forests
Biotic impacts
been about 0.85°C
(IPCC, 2013), with 2015 having been the first
year with temperatures more than 1°C above pre-industrial
Distribution of pests
temperatures (
Hawkins et al., 2017). Global meta-analyses have
One of the most important changes likely to result from climate
documented significant range-boundary changes for 279 spe-
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
change is a shift in suitable habitats for certain pests, which is
cies, which, on average, have shifted poleward by 40 km over an
mainly linked to changing temperatures. Temperature influences
average timespan of 66 years
(Parmesan and Yohe, 2003).
thresholds for pest growth and survival through events such as
frost frequency and the requirement for reproduction as deter-
mined through accumulation of thermal units. A benchmark for
Geographic source of future pests
the effects of temperature on changes in distribution is provided
by the relationship of temperature with elevation and latitude
The main pathways for the arrival of pests and pathogens are
(Linacre, 1992). Average warming over the past century has
associated with international trade. This section reviews recent
changes in trade patterns and considers future trends based on
trade agreements and expert opinion. New Zealand-specific
import data were obtained from
Statistics New Zealand (2017)
and for data going back to 1988, from
Statistics NZ Infoshare
(2017).
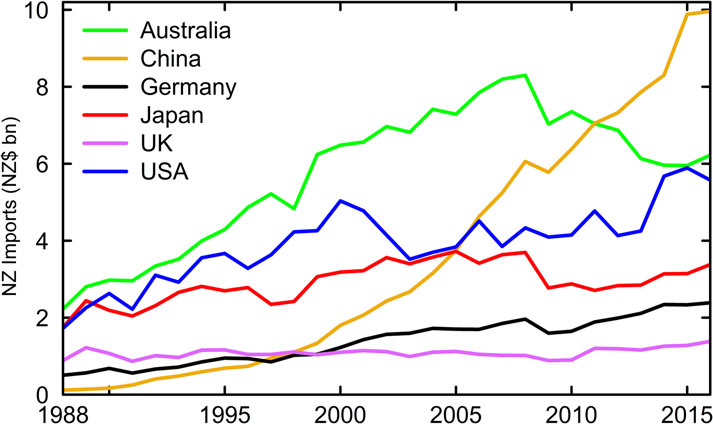
Since the 1980s, imports have increased from the estab-
lished trading countries of Australia, Europe, US and Japan.
However, the most significant change has been the dramatic
rise of China as the dominant importer to New Zealand
(Figure
7). In 1988, only 1 per cent of New Zealand’s imports ori-
ginated from China, but in 2016 these imports represented
more than 16 per cent.
Changes in the origins of imported goods and passengers are
expected to result in changes in the profile of pest threats. In fact,
this has already been observed in several regions. For example, in
North America and Europe there has been an increase in insect
pest and pathogen incursions originating from north-east Asia
(
Brockerhoff and Liebhold, 2017). Historically, the majority of forest
Figure 5 Multi-model-mean projections of annual frequency of Very
insects invading North America originated from Europe but in the
High and Extreme (VH + E) forest fire danger over fire season months
last two decades, north-east Asian species, including some high-
(Oct–Apr). Note the non-linear colour scale. The locations used to con-
impact invaders, such as the Asian Longhorned beetle (detected in
struct this figure are shown as black dots, and actual fire risks for these
1998) and Emerald Ash Borer (detected in 2002), have become
locations is given in Figure
6.
more prominent
(Aukema et al., 2010).
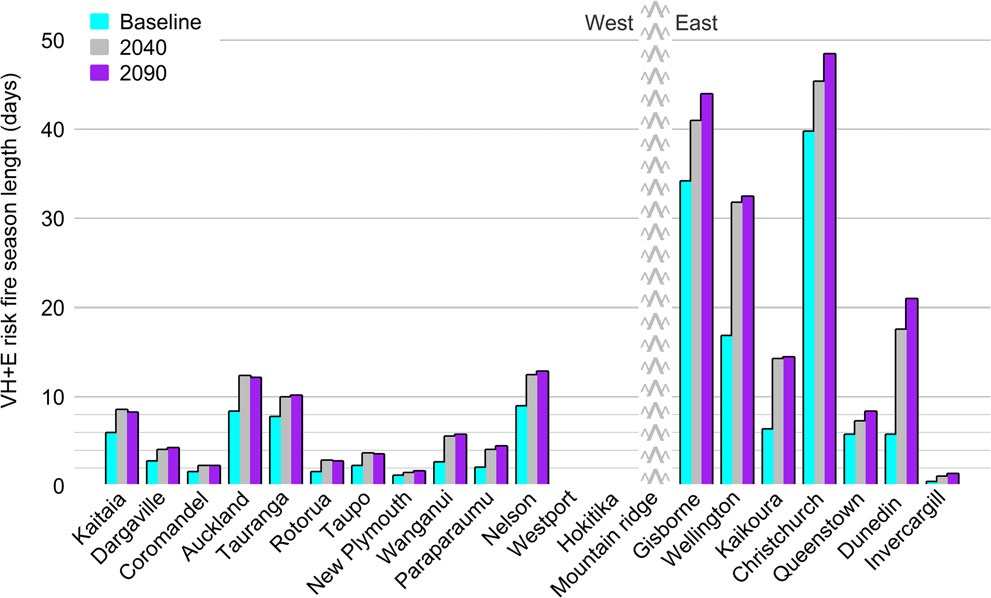
Figure 6 Projected average number of days of the fire season with Very High and Extreme (VH + E) forest fire danger under current conditions and
in 2040 and 2090, at individual station locations and averaged for New Zealand across the 12 GCMs. Locations were grouped with respect to New
Zealand’s main mountain ranges, then ordered by latitude (northern-most left).
7
link to page 13 link to page 14 link to page 14 link to page 14 link to page 14 link to page 12 link to page 13 link to page 8 link to page 15 link to page 15 link to page 13 link to page 9 link to page 13 link to page 12 link to page 14
Forestry
Table 3 Variation in mean predicted stand severity, Ssev, of cyclaneusma
needle cast and dothistroma needle blight for New Zealand under
current climate and the B1, A1B and A2 emission scenarios, projected
for 2040 and 2090 within the North (NI) and South (SI) Islands.
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
Year
Emission
Cyclaneusma
Dothistroma
scenario
NI
SI
NI
SI
Baseline
6.40
2.17
10.9
4.26
2040
B1
6.18
3.56
10.0
5.04
A1B
6.16
3.96
9.52
5.44
A2
6.17
3.85
9.54
5.47
2090
B1
6.03
4.76
8.69
6.13
A1B
5.62
5.76
7.51
6.83
Figure 7 Imports from New Zealand’s main trading partners, 1988-
A2
5.25
6.17
5.82
7.89
2016 (
Statistics New Zealand, 2017).
Based on import trends and bilateral/multilateral trade
Climate matching of current and future climates suggests that
agreements, one can assume that New Zealand’s imports from
parts of north-east Asia, southern South America, western
east Asia will continue to increase. New Zealand’s traditional
Europe, and southern Africa, as well as cooler (e.g. montane)
trading partners in Europe and North America are likely to
regions in subtropical countries may represent sources of spe-
remain important sources of imports, even though their relative
cies of growing biosecurity concern (
Ridley et al., 2000;
Peacock
share may decrease, and Australia is likely to remain a key
and Worner, 2006; Bertheau et al., 2010; Kriticos, 2012).
source of imports owing to its physical proximity. While trade
may arguably present the greatest risk of introducing unwanted
pests and diseases, the increasing number of international pas-
senger arrivals to New Zealand presents another important
Damage from tree pathogens
pathway for possible pest incursion.
Within the major plantation areas in the North Island, simula-
The volume of imports and the number of international visitors
tions showed that the severity of dothistroma needle blight and
may be primary drivers of pest propagule pressure, but several
cyclaneusma needle cast are likely to decline throughout the
other factors will also affect future biosecurity risks. For example,
21st century under all emission scenarios; however, increases in
rapidly growing trade with new trading partners may have a dis-
disease severity were predicted for large areas of the South
proportionate effect on biosecurity risks because they may host
Island (Table
3). With the exception of the west coast of the
pests that have not previously had access to New Zealand. Many
South Island, the actual predicted severity remains relatively
of the potentially most invasive species from New Zealand’s long-
low compared to the damaging levels currently found within
standing trading partners have either established themselves in
the North Island. Although high disease severity is predicted
New Zealand already or have been excluded by effective border
within the west coast of the South Island under both projection
biosecurity measures targeted at specific species or to mitigate
periods (2040 and 2090), it causes little concern in the national
specific entry pathways. Such measures include, for example,
context because this area currently constitutes only 1.8 per cent
requirements for pest monitoring, control and treatment by over-
of the total New Zealand plantation area
(Watt et al., 2011a,
seas growers and exporters, border inspection and additional
2012a).
treatment requirements for imports, and post-border pest surveil-
There are also a number of other pathogens that could cause
lance and incursion response capability (
Gordh and McKirdy, 2013).
considerable damage to plantation forests should they establish
Pest-specific measures are not necessarily in place for pests from
within New Zealand. Pitch canker is a devastating disease of
newer non-traditional trading partners, although many pests may
Pinus spp., and P. radiata is known to be highly susceptible to
be excluded by generic measures to prevent pest entry. The great-
the disease
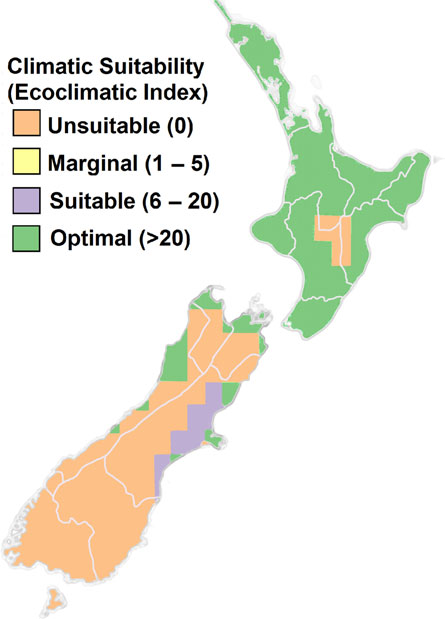
(Ganley et al., 2011). Projections using process-
est threat may be from new ‘hitchhiker’ species (i.e. species that
based distribution models show that the potential New Zealand
are transported inadvertently on inanimate objects such as sea
distribution of this disease could expand from coastal areas of
containers or vehicles) that are difficult to manage because they
the North Island under baseline climate to almost all of the
are not necessarily associated with particular pathways in a pre-
North Island and eastern parts of the South Island by 2080
dictable way (
Toy and Newfield, 2010).
(Figure
8) (
Ganley et al., 2011).
Climatic similarities between New Zealand and the potential
Red needle cast caused by Phytophthora pluvialis causes peri-
incursion species’ native habitat will also be important for the
odic episodes of defoliation in New Zealand’s P. radiata planta-
ability of pests and plant weeds to establish and develop pest
tions, particularly in locations exposed to frequent wet days and
potential in New Zealand. In this respect the southeast Asian
fog over the cooler months. Phytophthora pinifolia causes similar
countries will probably pose smaller risks due to their funda-
problems in Chile
(Durán et al., 2008) and could be problematic
mentally different climatic zones. However, pests from temper-
in New Zealand if it were introduced. Perhaps a greater threat is
ate and some subtropical regions are of greater concern.
posed by pathogens that are new to science (e.g. P. pluvialis was
8
link to page 12 link to page 12 link to page 12 link to page 13 link to page 14 link to page 13 link to page 14 link to page 13 link to page 13 link to page 13 link to page 12 link to page 12 link to page 12 link to page 12 link to page 12 link to page 12 link to page 15 link to page 13 link to page 13 link to page 13 link to page 14 link to page 13 link to page 14 link to page 14 link to page 14 link to page 13 link to page 14 link to page 13 link to page 13 link to page 13 link to page 13 link to page 12 link to page 12 link to page 12 link to page 13 link to page 13
Climate change effects on plantation forests
Zealand, most insects feeding on these trees are non-native
species that were introduced accidentally, but see
Berndt et al.
(2004) for a description of a native defoliator that has adapted
to feeding on pines. Fortunately, most of these species cause lit-
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
tle or no damage at present. An exception is the woodwasp
Sirex noctilio that was a concern in the past, but is now largely
controlled through the introduction of biological control agents
and changes in forest management that reduce stand suscepti-
bility to S. noctilio
(Bain et al., 2012). However, the pest status of
some of these species could change as a result of climate
change, and in other countries, there are many pests of conifers
that could represent serious threats to plantation forests in New
Zealand if they should ever become established in the country
(
Brockerhoff and Bulman, 2014). These threats could potentially
become even more severe in the future.
There are hundreds of damaging pests associated with conifers
that are not yet present in New Zealand
(Brockerhoff and Bulman,
2014). For example, the Eurasian nun moth, Lymantria monacha,
and the European pine processionary moth, Thaumetopoea pityo-
campa, would probably cause considerable defoliation if they
became established in New Zealand
(Withers and Keena, 2001;
Kriticos et al., 2013). A climate-matching study has indicated that
Figure 8 Pitch canker Ecoclimatic Index for 2080 derived from NCAR-
there are also a number of North American bark beetles that
CCSM for the A1B emissions scenario. Based on
Ganley et al., 2011. The
attack pines and could present threats to New Zealand’s planta-
squares evident in the figure reflect the resolution of the underlying cli-
tion forests. They include Dendroctonus valens and Ips calligraphus
mate change scenario.
(
Lantschner et al., 2017).
Most of the regions in New Zealand where conifer plantation
unknown when the pathogen was first isolated from diseased P.
forests occur are expected to be climatically suitable for the pine
radiata foliage in 2008), or by those that behave in unexpected
processionary moth, and estimates of its impacts on pine prod-
ways in a new environment. For instance, Neonectria fuckeliana
uctivity suggest that significant growth losses would occur if it
(C. Booth) Castl. & Rossman was well known in the Northern
were introduced
(Kriticos et al., 2013). In Europe, the main limit-
Hemisphere as a wound-invading fungus that was found only
ing factor for most insect pests is cold stress (
Robinet et al., 2007;
on spruce (Picea spp.) and fir (Abies spp.) and caused little or no
Kriticos et al., 2013). Hence, for many species, climate change is
damage. When it first established in the Southern Hemisphere
expected to increase the area with suitable climate
(Robinet
on P. radiata, it caused severe damage to some plantations in
et al., 2007). The situation in New Zealand is likely to be similar.
the lower half of the South Island (
Crane et al., 2009) until a
This effect of climate change on climatic suitability is likely to
successful control strategy was developed.
have more far-reaching implications for species from warmer
(e.g. subtropical) regions which currently are unlikely to become
established in New Zealand given its mainly temperate climate.
Damage from tree-feeding insects
Several studies have suggested that climate change will increase
Although climate change effects on tree-feeding insects are rela-
the risk of establishment of species from warm-temperate or
tively well known in several countries, and have been the subject
subtropical regions
(Peacock and Worner, 2006; Kriticos, 2012).
of a considerable research effort, little research on this topic has
Climate change may also affect the severity of damage from
been carried out in New Zealand. However, several general
existing insect pests because warmer temperatures can be
reviews specific to New Zealand exist, and one specific study on
expected to accelerate insect development and therefore lead
effects by a potentially invasive defoliator was undertaken.
to an increase in population levels, especially in species that can
Climate change can affect problems related to insect pests
complete more than one generation per year. An example of
through several mechanisms including (i) changing the severity
such a species is the Monterey pine aphid, Essigella californica
of damage by native or non-native insects due to changes in cli-
(
Watson et al., 2008). Although this aphid is presently not con-
matic suitability for the pest or the host tree
(Battisti et al.,
sidered damaging in New Zealand, in parts of Australia with a
2005;
Marini et al., 2012), (ii) changing the likelihood of estab-
warmer climate (i.e. warmer than New Zealand’s current cli-
lishment of invasive species that are not yet present in New
mate), E. californica can cause considerable defoliation of pines
Zealand (
Sutherst et al., 2007;
Kriticos et al., 2013), and (iii) as
(
May and Carlyle, 2003).
an indirect consequence through interactions with other disturb-
In other parts of the world, warming has been shown to
ance factors, such as increases in fire hazards due to tree mor-
increase population levels and damage by the mountain pine
tality or pest susceptibility of stands through increasing wind
beetle and other bark beetles
(Hicke et al., 2006;
Marini et al.,
damage (
Stinson et al., 2011;
Hickey et al., 2012;
Jenkins et al.,
2012;
Bentz and Jönsson, 2015;
Bentz et al., 2016). Warming
2012).
can thus lead to an expansion of areas affected by tree-feeding
Currently, only a few insect pests affect P. radiata plantations
insects
(Battisti et al., 2005; Marini et al., 2012). The unprece-
in New Zealand. As there are no native Pinaceae in New
dented spread of the mountain pine beetle into the boreal
9
link to page 12 link to page 12 link to page 12 link to page 13 link to page 13 link to page 14 link to page 14 link to page 14 link to page 14 link to page 14 link to page 15 link to page 15 link to page 13 link to page 15 link to page 12 link to page 15 link to page 15 link to page 14 link to page 14 link to page 14 link to page 14 link to page 13 link to page 14 link to page 15 link to page 15 link to page 15 link to page 15 link to page 15 link to page 14 link to page 14 link to page 15 link to page 14 link to page 13 link to page 14 link to page 13 link to page 13 link to page 13 link to page 13
Forestry
forest east of the Rocky Mountains, as a result of climate
New Zealand. Although we do not currently have an estimate
change, is of particular concern because it could cause substan-
of the potential distribution of this species, the distribution
tial mortality of jack pine, Pinus banksiana Lamb., and other
where it is invasive in the US is quite similar to that of M. quin-
eastern pines (
Cullingham et al., 2011). Furthermore, warmer
quenervia. During the 1990s, kudzu has migrated northwards
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
temperatures, especially if associated with greater frequency of
in the continental US from its original distribution, a shift which
drought conditions, can increase the susceptibility of trees to
is in line with previous model predictions
(Sasek and Strain,
attack and damage from bark beetles or wood borers such as S.
1990). This change demonstrates the responsiveness of the
noctilio
(Carnegie and Bashford, 2012). Both warming and
weed to climatic conditions and highlights that the potential
increasing drought incidence can lead to an expansion of areas
for range expansion under climate change should not be
affected by tree-feeding insects (
Battisti et al., 2005; Marini
underestimated.
et al., 2012). Although no identified insect pest in plantation for-
There is a risk that currently established exotic woody tree
ests in New Zealand has yet been observed to have increased in
species native to Australia may become more dominant compe-
its severity through climate change, this is likely to occur in the
titors in New Zealand under a warmer climate. Acacia spp. have
future.
very high growth rates and can rapidly occupy disturbed sites,
vigorously competing with planted P. radiata seedlings (
Turvey
et al., 1983). As tree species, they can compete further into the
rotation than even tall weed species
(Hunt et al., 2006), which
Competition with weeds
are predominantly shrubs. Some species have the ability to
The future prevalence of weed problems is likely to be related to
resprout after their stems have been severed which makes
(i) the future distribution, growth and competitive strength of
them hard to control. Seed germination is also often stimulated
the currently problematic weeds, and (ii) the potential of weeds
by fire. The likely increases in fire frequency and severity will
that are either already present in New Zealand, but not yet
make sites more predisposed to invasion by these species. Some
widely distributed, or that could enter the country to become
Acacia species are already a localized problem in northern and
problem weeds, especially under changed climatic conditions.
eastern parts of the country (
Watt et al., 2008).
The weed species that compete most strongly with P. radiata
Climate change is also likely to affect growth rates of weeds
within New Zealand are tall woody weeds. Within this group,
through changes in CO2, root-zone water storage, temperature
gorse (Ulex europaeus), Scotch broom (Cytisus scoparius), brack-
and changing length of the growing season. If relative growth
en (Pteridium esculentum), blackberry (Rubus fruticosus) and
of both plantation trees and weeds increases at the same rate
wilding conifers (e.g. Pinus contorta and Pseudotsuga menziesii)
then competition levels may not significantly change. However,
are the most competitive and invasive (
Watt et al., 2008). In
in agricultural settings there is evidence that weeds exhibit a
addition, buddleja (Buddleja davidii), Acacia spp. and pampas
stronger positive response to CO2 than crop plants which is likely
(Cortaderia spp.) are problematic weeds in some specific regions.
to lead to reductions in crop yields
(Ziska et al., 2011;
Ziska,
Tall shrubby species reduce plantation growth more than short
2011). The basis for this increased competitive behaviour of
species such as grasses and herbaceous species, as they com-
weeds is unclear but may be related to the vigorous and gener-
pete more vigorously for both water and light and are not as
ally indeterminate growth habit of weeds and greater genetic
effectively shaded out as trees grow taller (
Richardson et al.,
and phenotypic plasticity associated with wild species (
Ziska
1999;
Watt, 2003).
and McConnell, 2015). The growth response of different weeds
Little research has investigated the future distribution of the
to climate change has been shown to vary widely
(Sheppard
most problematic weed species in New Zealand under climate
and Stanley, 2014). As a consequence, increases in CO2 have
change.
Potter et al. (2009) found that changes in climate will
been shown to preferentially select for more responsive invasive
have little effect on the potential distribution of broom, with all
species within plant communities (
Ziska and McConnell, 2015).
regions remaining suitable for the species. In contrast, it is
expected that under future climate change, buddleja may
expand its range within the southeast of New Zealand (
Watt
Discussion
et al., 2010;
Kriticos et al., 2011).
Expansion of ‘sleeper weeds’ is likely to pose a future threat to
Climate change impact assessments of forest systems are often
plantation forests. Sleeper weeds are weeds that are present in
dominated by a focus on biophysical factors, which generally
New Zealand, but whose distribution or vigour is limited under
show positive effects of climate change on forest productivity
current climatic conditions. For instance, the exotic tree Melaleuca
(
Reyer, 2015). This is largely due to the growth response to ele-
quinquenervia, which is currently established in Auckland and
vated CO2
(Hickler et al., 2015; Reyer, 2015), and even tempera-
Northland, could become quite invasive if the species’ thermal
ture increases by a few degrees, or precipitation changes by a
requirement for reproduction within northern areas of New
few per cent, especially when coupled with increasing CO2 con-
Zealand is surpassed in the future
(Watt et al., 2009). Range
centrations, may not be of great detriment to the physiological
expansion of this species could have significant consequences as
growth potential of many forest stands (e.g.
Kirschbaum et al.,
M. quinquenervia has been found to be extremely difficult to con-
2012).
trol in exotic locations, such as Florida (
Austin, 1978; Woodall,
Other factors impinging on the fate of forest stands, on the
1983).
other hand, can be much more negative. This has been experi-
Kudzu (Pueraria montana) is a perennial, semi-woody, climb-
enced most clearly by the expansion of the mountain pine bee-
ing leguminous vine, which is extremely invasive and damaging
tle in North America (
Hicke et al., 2006;
Marini et al., 2012) that
in the south-eastern US. It has recently been found in northern
has the potential to lead to the death of susceptible stands that
10
link to page 12 link to page 13 link to page 12 link to page 14 link to page 13 link to page 13 link to page 12 link to page 13 link to page 12 link to page 12 link to page 12 link to page 13 link to page 14 link to page 14 link to page 12 link to page 13 link to page 13
Climate change effects on plantation forests
had previously been protected by mortality of the insect pests
circumstances, such as under severely nutrient-limited condi-
during severe winters
(Cullingham et al., 2011). Because of the
tions (e.g.
Ceulemans and Mousseau, 1994;
Kirschbaum, 2011).
tight linkage between temperature and distribution of the pest
Empirical evidence in support of these various positions is mixed
species, its range expansion under future warming can be antici-
(e.g.
Norby et al., 1999; Nowak et al., 2004;
Donohue et al., 2013;
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
pated with very high probability. This pattern is expected to pre-
Kirschbaum and Lambie, 2015), but a recent reanalysis of the
dominate for many pest species (e.g.
Logan et al., 2003).
results of past short-term CO2 growth experiments suggested
Similarly, wildfire risk is strongly linked to climatic factors
that future growth enhancement may lie about half way
such as temperature, humidity and wind speed which, together
between the values calculated with constant and increasing
with stand attributes like fuel load and fuel moisture contents,
CO2, respectively
(Kirschbaum and Lambie, 2015). Clearly, more
largely control fire risks in forest systems. These factors are gen-
research is still needed to refine the likely growth response to
erally expected to change towards increasing fire risk in the
increasing CO2 of plantations in New Zealand and elsewhere.
future (e.g.
Flannigan et al., 2009;
Pechony and Shindell, 2010).
That single factor constitutes the largest single uncertainty in
Forest stands are also at risk from wind damage, although cli-
current modelling of plant responses to future conditions and
mate change risks for wind are more nuanced and depend on
makes it still difficult to confidently forecast changes to prod-
changes in climatic factors as much as changes in stand proper-
uctivity and risk of wind damage.
ties (
Moore and Watt, 2015).
Current projections show that the risks to plantations from
For a balanced assessment of the combined climate change
the two most damaging diseases in New Zealand are unlikely to
effects of all agents of change, it is therefore necessary to con-
change markedly. Nonetheless, further research should be
sider all of the key aspects of change in an integrated assess-
undertaken to examine the potential impact of the damaging
ment. This requires collaboration between different science
disease red needle cast (P. pluvialis) under current and future cli-
disciplines and the bringing-together of different approaches
mates. Currently, there is also no significant damage to planta-
and numeric evaluations. The present work took a first step in
tions from insect pests. However, it would be useful to
such an assessment by cataloguing the various changes that
determine how climate change affects the climatic suitability of
our forests might be subject to in the future. A further stage of
plantation forest regions for a range of high-risk species such as
development in an integrated assessment would see the differ-
bark beetles and defoliators. The impact of weeds on planta-
ent processes combined in unified ecosystem models that can
tions in the future is likely to depend on the degree to which
also quantitatively integrate these processes. Although linkages
current ‘sleeper’ weeds and naturalised aggressive woody tree
were included between predictions of productivity and wind
species (e.g. Acacia spp.) can expand their range and increase in
damage, most effects that were assessed in this study were not
vigour. The future potential distributions of these key weed spe-
linked in a dynamic way which is a limitation of our approach.
cies should be determined using process-based weed distribu-
For instance, by direct inclusion of pest and disease damage in
tion models. Further research is also required to determine how
a comprehensive model, the plant physiological status could be
important weed species will respond to climate change and
directly linked to plant susceptibility to certain diseases.
how effectively they will compete with plantations in the future.
Conversely, any pest or disease damage could directly affect
Given the potentially damaging role wind and fire are projected
plant growth, with consequences for future photosynthetic car-
to have on future plantations, it would also be useful to refine
bon gain or, vulnerability to wind or fire damage. However, this
our spatial understanding of the impacts of these factors.
would require significantly greater system complexity and math-
ematical integration of very different model components. Such
integration will ultimately be needed for a true assessment of
Conclusion
climate change impacts and avoidance of any bias in climate
impact assessments brought about by omission of any import-
Overall, our analyses showed productivity gains for P. radiata
ant aspects of system processes and interactions.
from the direct effects of climate change that ranged from rela-
tively minor to substantial depending on the response to
increasing CO2. These simulations suggest that the direct effects
Sensitivities and areas for future research
of climate change are likely to favourably affect forest product-
ivity even if the potential CO2 response is only partly realized.
This review identified the growth response to increasing CO2 as
Although fire risk is projected to increase in the future, most
a key sensitivity of the overall response of forest systems to
damage is likely to ensue from the greater vulnerability of plan-
future climate change, but there is still considerable uncertainty
tations to wind damage, that results from increased height
of the magnitude of this response. Some workers have focused
growth.
on photosynthetic processes and expect large increases in CO2
Changes in trade and increased global travel are likely to
responsiveness, especially under water-limited conditions (e.g.
influence the origin of future incursions of invasive pests, with
Lloyd and Farquhar, 1996; Franks et al., 2013). Others have
invasions from regions in eastern Asia likely to constitute a
argued that stimulation of photosynthesis is inconsequential
growing risk. Currently, the most significant biotic disturbances
under many natural conditions and can be overridden by other
of New Zealand plantations come from two needle cast dis-
growth-limiting factors (e.g.
Körner et al., 2007; Fatichi et al.,
eases, for which climate projections show very little change in
2014). Others have presented a more diverse picture, suggesting
damage over the course of this century. Although New Zealand
that CO2 stimulated carbohydrate supply may stimulate growth
does not currently have any damaging forest insect species,
under some conditions, especially under low light or water
population levels and resulting damage are likely to increase in
stress, but may have little effect on productivity in other
the
future
as
warmer
temperatures
accelerate
insect
11
link to page 7
Forestry
development and increase the susceptibility of host plants to
moth caused by increased winter temperatures. Ecol. Appl. 15,
attack. Competition within plantations from aggressive woody
2084–2096.
tree species, and in particular those originating from Australia is
Bentz, B.J., Duncan, J.P. and Powell, J.A. 2016 Elevational shifts in ther-
likely to increase as a result of climate change.
mal suitability for mountain pine beetle population growth in a changing
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
The effects of climate change present global plantation for-
climate. Forestry 89, 271–283.
ests with many challenges but also new opportunities. This
Bentz, B.J. and Jönsson, A.M. 2015 Modeling bark beetle responses to cli-
study quantifies the increases in productivity expected due to
mate change. In Bark Beetles: Biology and Ecology of Native and Invasive
climate change. Changes in wind conditions will have implica-
Species. F.E. Vega and R.W. Hofstetter (eds). Academic Press, pp.
tions for silvicultural practices, particularly for stands grown to
533–553.
maximize carbon, while there will be significantly increased wild-
Berndt, L., Brockerhoff, E.G., Jactel, H., Weiss, T. and Beaton, J. 2004
fire risk to plantation forests. The future impact from biotic fac- Biology and rearing of Pseudocoremia suavis, an endemic looper
tors are complex and often species dependent, but this study
(Lepidoptera: Geometridae) with a history of outbreaks on exotic coni-
highlights the major threat species and notes the highest-risk
fers. N. Z. Entomol. 27, 73–82.
source locations. The results from this ambitious synthesis of cli-
Bertheau, C., Brockerhoff, E.G., Roux‐Morabito, G., Lieutier, F. and Jactel,
mate change threats should provide decision makers the fore-
H. 2010 Novel insect‐tree associations resulting from accidental and
sight to mitigate against avoidable threats, adapt to committed
intentional biological ‘invasions’: a meta‐analysis of effects on insect fit-
future changes and capitalize on future opportunities.
ness. Ecol. Lett. 13, 506–515.
Betts, R.A., Jones, C.D., Knight, J.R., Keeling, R.F. and Kennedy, J.J. 2016
El Nino and a record CO2 rise. Nature Clim. Change 6, 806–810.
Acknowledgements
Brockerhoff, E. and Bulman, L. 2014 Biosecurity risks to New Zealand’s
plantation forests and the rationale for pathway risk management. N. Z.
We wish to thank Duncan Harrison of Scion for implementing the co-
J. Forestry 59, 3–8.
kriging technique and re-creating the maps in Figure
5. We are very
grateful to the Chief Editor, Associate Editor and two anonymous
Brockerhoff, E. and Liebhold, A. 2017 Ecology of forest insect invasions.
reviewers for their useful suggestions which greatly improved the
Biol. Invasions 19, 3141–3159.
manuscript.
Cameron, A.D. 2002 Importance of early selective thinning in the devel-
opment of long-term stand stability and improved log quality: a review.
Forestry 75, 25–35.
Funding
Carnegie, A.J. and Bashford, R. 2012 Sirex woodwasp in Australia: current
New Zealand Ministry for Primary Industries through the Sustainable
management strategies, research and emerging issues. In The Sirex
Land
Management
and
Climate
Change
(SLMACC)
programme.
Woodwasp and its Fungal Symbiont. B. Slippers, P. De Groot and M.J.
Additional support via MBIE core funding (C04X1104) to Scion.
Wingfield (eds). Springer, pp. 175–201.
Ceulemans, R. and Mousseau, M. 1994 Tansley review no. 71 effects of
elevated atmospheric co2on woody plants. New Phytolog. 127, 425–446.
Conflict of interest statement
Chen, W., Chen, J. and Cihlar, J. 2000 An integrated terrestrial ecosystem
None declared.
carbon-budget model based on changes in disturbance, climate, and
atmospheric chemistry. Ecol. Modell. 135, 55–79.
Cramer, W., Yohe, G., Auffhammer, M., Huggel, C., Molau, U., Dias, M.S.,
et al. 2014 Detection and attribution of observed impacts. In Climate
References
Change 2014: Impacts, Adaptation, and Vulnerability. Cambridge
Allen, C.D., Macalady, A.K., Chenchouni, H., Bachelet, D., McDowell, N.,
University Press, pp. 979–1038.
Vennetier, M., et al. 2010 A global overview of drought and heat-induced
Crane, P.E., Hopkins, A.J.M., Dick, M.A. and Bulman, L.S. 2009 Behaviour
tree mortality reveals emerging climate change risks for forests. For.
of Neonectria fuckeliana causing a pine canker disease in New Zealand.
Ecol. Manage. 259, 660–684.
Can. J. For. Res. 39, 2119–2128.
Anderegg, W.R., Hicke, J.A., Fisher, R.A., Allen, C.D., Aukema, J., Bentz, B.,
Cullingham, C.I., Cooke, J.E.K., Dang, S., Davis, C.S., Cooke, B.J. and
et al. 2015 Tree mortality from drought, insects, and their interactions in
Coltman, D.W. 2011 Mountain pine beetle host-range expansion threa-
a changing climate. New Phytol. 208, 674–683.
tens the boreal forest. Mol. Ecol. 20, 2157–2171.
Anderson, S. 2005 Forest and rural fire danger rating in New Zealand. In:
Donohue, R.J., Roderick, M.L., McVicar, T.R. and Farquhar, G.D. 2013
Forestry Handbook. M. Colley (ed.). New Zealand Institute of Forestry. pp
Impact of CO2 fertilization on maximum foliage cover across the globe’s
241–244.
warm, arid environments. Geophys. Res. Lett. 40, 3031–3035.
Aukema, J.E., McCullough, D.G., Von Holle, B., Liebhold, A.M., Britton, K.
Durán, A., Gryzenhout, M., Slippers, B., Ahumada, R., Rotella, A., Flores, F.,
and Frankel, S.J. 2010 Historical accumulation of nonindigenous forest
et al. 2008 Phytophthora pinifolia sp. nov. associated with a serious nee-
pests in the continental United States. Bioscience 60, 886–897.
dle disease of Pinus radiata in Chile. Plant Pathol. 57, 715–727.
Austin, D.F. 1978 Exotic plants and their effects in southeastern Florida.
Fatichi, S., Leuzinger, S. and Körner, C. 2014 Moving beyond photosyn-
Environ. Conserv. 5, 25–34.
thesis: from carbon source to sink‐driven vegetation modeling. New
Bain, J., Sopow, S.L. and Bulman, L.S. 2012 The Sirex woodwasp in New
Phytol. 201, 1086–1095.
Zealand: history and current status. In The Sirex Woodwasp and its
Flannigan, M.D., Krawchuk, M.A., de Groot, W.J., Wotton, B.M. and
Fungal Symbiont. B. Slippers, P. De Groot and M.J. Wingfield (eds).
Gowman, L.M. 2009 Implications of changing climate for global wildland
Springer, pp. 167–173.
fire. Int. J. Wldland fire 18, 483–507.
Battisti, A., Stastny, M., Netherer, S., Robinet, C., Schopf, A., Roques, A.,
Franks, P.J., Adams, M.A., Amthor, J.S., Barbour, M.M., Berry, J.A.,
et al. 2005 Expansion of geographic range in the pine processionary
Ellsworth, D.S., et al. 2013 Sensitivity of plants to changing atmospheric
12
Climate change effects on plantation forests
CO2 concentration: from the geological past to the next century. New
Kirschbaum, M.U.F. and Watt, M.S. 2011 Use of a process-based model
Phytol. 197, 1077–1094.
to describe spatial variation in Pinus radiata productivity in New
Ganley, R., Watt, M., Kriticos, D., Hopkins, A. and Manning, L. 2011
Zealand. For Ecol. Manage. 262, 1008–1019.
Increased risk of pitch canker to Australasia under climate change.
Kirschbaum, M.U.F., Watt, M.S., Tait, A. and Ausseil, A.G.E. 2012 Future
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
Australas Plant Pathol. 40, 228–237.
wood productivity of Pinus radiata in New Zealand under expected cli-
Gardiner, B.A., Peltola, H. and Kellomaki, S. 2000 Comparison of two
matic changes. Glob Change Biol 18, 1342–1356.
models for predicting the critical wind speeds required to damage con-
Kriticos, D.J. 2012 Regional climate-matching to estimate current and
iferous trees. Ecol. Modell. 129, 1–23.
future sources of biosecurity threats. Biol. Invasions 14, 1533–1544.
Goovaerts, P. 1999 Using elevation to aid the geostatistical mapping of
Kriticos, D.J., Leriche, A., Palmer, D.J., Cook, D.C., Brockerhoff, E.G.,
rainfall erosivity. Catena 34, 227–242.
Stephens, A.E.A., et al. 2013 Linking climate suitability, spread rates and
Gordh, G. and McKirdy, S. 2013 The Handbook of Plant Biosecurity:
host-impact when estimating the potential costs of invasive pests. PLoS
Principles and Practices for the Identification, Containment and Control of
One 8, e54861.
Organisms that Threaten Agriculture and the Environment Globally.
Kriticos, D., Watt, M., Potter, K., Manning, L., Alexander, N. and Tallent-
Springer Science & Business Media.
Halsell, N. 2011 Managing invasive weeds under climate change: consid-
Goulding, C.J. 1994 Development of growth models for Pinus radiata in
ering the current and potential future distribution of Buddleja davidii.
New Zealand – experience with management and process models. For.
Weed Res. 51, 85–96.
Ecol. Manage. 69, 331–343.
Kurz, W.A., Dymond, C., Stinson, G., Rampley, G., Neilson, E., Carroll, A.,
Hawkins, E., Ortega, P., Suckling, E., Schurer, A., Hegerl, G., Jones, P., et al.
et al. 2008 Mountain pine beetle and forest carbon feedback to climate
2017 Estimating changes in global temperature since the pre-industrial
change. Nature 452, 987–990.
period. Bull. Am. Meteorolog. Soc. 98, 1841–1856.
Körner, C., Morgan, J. and Norby, R. 2007 CO2 fertilization: when, where,
Hawkins, E. and Sutton, R. 2009 The potential to narrow uncertainty
how much? In Terrestrical Ecosystems in a Changing World. Canadell J.G.,
in regional climate predictions. Bull. Am. Meteorolog. Soc. 90,
Pataki D.E. and Pitelka L.F. (eds). Springer Verlag, pp. 9–21.
1095–1107.
Lantschner, M.V., Atkinson, T.H., Corley, J.C. and Liebhold, A.M. 2017
Hawkins, E. and Sutton, R. 2011 The potential to narrow uncertainty in
Predicting North American Scolytinae invasions in the Southern
projections of regional precipitation change. Clim. Dyn. 37, 407–418.
Hemisphere. Ecol. Appl. 27, 66–77.
Hicke, J.A., Logan, J.A., Powell, J. and Ojima, D.S. 2006 Changing tem-
Linacre, E. 1992 Climate Data and Resources: A Reference and Guide.
peratures influence suitability for modeled mountain pine beetle
Psychology Press.
(Dendroctonus ponderosae) outbreaks in the western United States. J.
Lloyd, J. and Farquhar, G. 1996 The CO2 dependence of photosynthesis,
Geophys. Res.—Biogeosci. 111, G02019.
plant growth responses to elevated atmospheric CO2 concentrations
Hickey, J.A., Johnson, M.C., Hayes, J.L. and Preisler, H.K. 2012 Effects of bark
and their interaction with soil nutrient status. I. General principles and
beetle-caused tree mortality on wildfire. For. Ecol. Manage 271, 81–90.
forest ecosystems. Funct. Ecol. 10, 4–32.
Hickler, T., Rammig, A. and Werner, C. 2015 Modelling CO2 impacts on
Logan, J.A., Régnière, J. and Powell, J.A. 2003 Assessing the impacts of
forest productivity. Curr. Forestry Rep. 1, 69–80.
global warming on forest pest dynamics. Front. Ecol. Environ. 1, 130–137.
Hunt, M.A., Battaglia, M., Davidson, N.J. and Unwin, G.L. 2006
Marini, L., Ayres, M.P., Battisti, A. and Faccoli, M. 2012 Climate affects
Competition between plantation Eucalyptus nitens and Acacia dealbata
severity and altitudinal distribution of outbreaks in an eruptive bark bee-
weeds in northeastern Tasmania. For. Ecol. Manage 233, 260–274.
tle. Clim. Change 115, 327–341.
IPCC 2013 Climate Change 2013: The Physical Science Basis. Contribution
May, B. and Carlyle, J. 2003 Effect of defoliation associated with
of
Working
Group
I
to
the
Fifth
Assessment
Report
of
the
Essigella californica on growth of mid-rotation Pinus radiata. For. Ecol.
Intergovernmental Panel on Climate Change. Cambridge University Press.
Manage, 183, 297–312.
Jenkins, M.J., Page, W.G., Hebertson, E.G. and Alexander, M.E. 2012 Fuels
Meehl, G.A., Stocker, T.F., Collins, W.D., Friedlingstein, P., Gaye, A.T.,
and fire behavior dynamics in bark beetle attacked forests in Western
Gregory, J.M., et al. 2007 Global Climate Projections. In Climate Change
North America and implications for fire management. For. Ecol. Manage
2007: The Physical Science Basis. Contribution of Working Group I to the
275, 23–34.
Fourth Assessment Report of the Intergovernmental Panel on Climate
Change. Solomon S., Qin D., Manning M., Chen Z., Marquis M., Averyt K.B.,
Kalnay, E., Kanamitsu, M., Kistler, R., Collins, W., Deaven, D., Gandin, L.,
et al. 1996 The NCEP/NCAR 40-year reanalysis project. Bull. Am.
Tignor M. and Miller H.L. (eds). Cambridge University Press, pp. 747–846.
Meteorol. Soc. 77, 437–472.
Melia, N., Haines, K. and Hawkins, E. 2015 Improved Arctic sea ice thick-
ness projections using bias-corrected CMIP5 simulations. Cryosphere 9,
Kean, J.M., Brockerhoff, E.G., Fowler, S., Gerard, P.J., Logan, D.P., Mullan,
A.B., et al. 2015 Effects of climate change on current and potential bio-
2237–2251.
security pests and diseases in New Zealand. Report for New Zealand
Ministry for the Environment 2008 Climate Change Effects and Impacts
Ministry for Primary Industries, MPI Technical Paper No: 2015/25.
Assessment. A Guidance Manual for Local Government in New Zealand.
Kirschbaum, M.U.F. 1999 CenW, a forest growth model with linked car-
2nd edn. Prepared for the Climate Change Office by B. Mullan, D. Wratt,
bon, energy, nutrient and water cycles. Ecol. Model 118, 17–59.
S. Dean, M. Hollis, S. Allan, T. Williams, G. Kenny and MfE. MFE Publication,
p. 149.
Kirschbaum, M. 2011 Does enhanced photosynthesis enhance growth?
Lessons learned from CO 2 enrichment studies. Plant Physiol. 155,
Mitchell, S., 2000. Forest health: preliminary interpretations for wind
117–124.
damage. In, Stand Density Management Diagrams. Forest Practices
Branch, B.C. Ministry of Forests, Victoria, B.C., p. 29.
Kirschbaum, M.U. and Lambie, S.M. 2015 Re-analysis of plant CO2
responses during the exponential growth phase: interactions with light,
Moore, J.R. and Watt, M.S. 2015 Modelling the influence of predicted
temperature, nutrients and water availability. Funct. Plant Biol. 42,
future climate change on the risk of wind damage within New Zealand’s
989–1000.
planted forests. Glob Change Biol 21, 3021–3035.
13
Forestry
Mullan, B., Caray-Smith, T., Griffiths, G. and Sood, A., 2011. Scenarios of
Sasek, T.W. and Strain, B.R. 1990 Implications of atmospheric CO2
storminess and regional wind extremes under climate change. NIWA
enrichment and climatic change for the geographical distribution of two
Client Report WLG2010-31 prepared for the Ministry of Agriculture and
introduced vines in the USA. Clim. Change 16, 31–51.
Forestry.
Scott, P. and Williams, N. 2014 Phytophthora diseases in New Zealand
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
Mullan, A.B., Wratt, D.S. and Renwick, J.A. 2002 Transient model scen-
forests. N. Z. J. Forestry 59, 15.
arios of climate changes for New Zealand. Weather Clim. 21, 3–34.
Seidl, R., Schelhaas, M.-J., Rammer, W. and Verkerk, P.J. 2014 Increasing
Norby, R.J., Wullschleger, S.D., Gunderson, C.A., Johnson, D.W. and
forest disturbances in Europe and their impact on carbon storage. Nat.
Ceulemans, R. 1999 Tree responses to rising CO2 in field experi-
Clim. Change 4, 806–810.
ments: implications for the future forest. Plant Cell Environ. 22,
Shanley, C.S., Pyare, S., Goldstein, M.I., Alaback, P.B., Albert, D.M., Beier, C.
683–714.
M., et al. 2015 Climate change implications in the northern coastal tem-
Nowak, R.S., Ellsworth, D.S. and Smith, S.D. 2004 Functional responses of
perate rainforest of North America. Clim. Change 130, 155–170.
plants to elevated atmospheric CO2 - do photosynthetic and productivity
Sheppard, C.S. and Stanley, M.C. 2014 Does elevated temperature and
data from FACE experiments support early predictions? New Phytol. 162,
doubled CO2 increase growth of three potentially invasive plants?
253–280.
Invasive Plant Sci. Manage. 7, 237–246.
NZFOA. 2016. Facts and Figures 2015/2016. New Zealand Forest Owners
Simpson, C.C., Pearce, H.G., Sturman, A.P. and Zawar-Reza, P. 2014
Association, 62 pp.
Verification of WRF modelled fire weather in the 2009–10 New Zealand
Parmesan, C. and Yohe, G. 2003 A globally coherent fingerprint of cli-
fire season. Int. J. Wildland Fire 23, 34–45.
mate change impacts across natural systems. Nature 421, 37–42.
Statistics, N.Z. Infoshare, 2017. Infoshare: Harmonised Trade – Imports,
Payn, T., Carnus, J.-M., Freer-Smith, P., Kimberley, M., Kollert, W., Liu, S.,
see
http://www.stats.govt.nz/infoshare/Default.aspx, accessed 15 May
et al. 2015 Changes in planted forests and future global implications.
2017.
For. Ecol. Manage. 352, 57–67.
Statistics New Zealandik, 2017. Global New Zealand – International
Peacock, L. and Worner, S. 2006 Using analogous climates and global
trade, investment, and travel profile: Year ended December 2017.
insect distribution data to identify potential sources of new invasive
Wellington: Ministry of Foreign Affairs and Trade, and Statistics New
insect pests in New Zealand. N. Z. J. Zool. 33, 141–145.
Zealand.
http://www.stats.govt.nz/browse_for_stats/industry_sectors/
Pearce, H.G., Kerr, J., Clark, A., Mullan, B., Ackerley, D., Carey-Smith, T.,
imports_and_exports/GoodsServicesTradeCountry_HOTPYeDec16/Related
et al., 2011. Improved estimates of the effect of climate change on NZ
%20Links.aspx, accessed on 15 May 2017.
fire danger. MAF Technical Paper No. 2011/13. Ministry of Agriculture –
Stinson, G., Kurz, W.A., Smyth, C.E., Neilson, E.T., Dymond, C.C.,
Forestry, Wellington. 84 pp.
Metsaranta, J.M., et al. 2011 An inventory-based analysis of Canada’s
Pechony, O. and Shindell, D.T. 2010 Driving forces of global wildfires over
managed forest carbon dynamics, 1990 to 2008. Glob. Change Biol. 17,
the past millennium and the forthcoming century. Proc. Natl. Acad. Sci.
2227–2244.
107, 19167–19170.
Sutherst, R.W., Baker, R.H.A., Coakley, S.M., Harrington, R., Kriticos, R. and
Potter, K.J.B., Kriticos, D.J., Watt, M.S. and Leriche, A. 2009 The current
Scherm, H., D.J., 2007. Pests Under Global Change - Meeting Your Future
and future potential distribution of Cytisus scoparius: a weed of pastoral
Landlords? In, Terrestrial Ecosystems in a Changing World. Springer,
systems, natural ecosystems and plantation forestry. Weed Res. 49,
Berlin, Heidelberg, New York, pp. 211-223.
271–282.
Tait, A. 2008 Future projections of growing degree days and frost in New
Quine, C.P. and Gardiner, B.A. 2002 Climate change impacts: storms. In
Zealand and some implications for grape growing. Weather Clim. 28,
Climate Change: Impacts on UK Forests. Broadmeadow M. (ed). Forestry
17–36.
Commission Bulletin, 125, HMSO, pp. 41–52.
Tait, A., Henderson, R., Turner, R. and Zheng, Z. 2006 Thin plate smooth-
Reeser, P., Sutton, W. and Hansen, E. 2013 Phytophthora pluvialis, a new
ing interpolation of daily rainfall for New Zealand using a climatological
species from mixed tanoak-Douglas-fir forests of western Oregon, USA.
rainfall surface. Int. J. Climatol. 26, 2097–2115.
North Am. Fungi 8, 1–8.
Tait, A. and Liley, B. 2009 Interpolation of daily solar radiation for New
Reyer, C. 2015 Forest productivity under environmental change – a
Zealand using a satellite derived cloud cover surface. Weather Clim. 29,
review of stand-scale modeling studies. Curr. Forestry Rep. 1, 53–68.
70–88.
Reyer, C.P., Bathgate, S., Blennow, K., Borges, J.G., Bugmann, H., Delzon,
Toy, S. and Newfield, M. 2010 The accidental introduction of invasive ani-
S., et al. 2017 Are forest disturbances amplifying or canceling out cli-
mals as hitchhikers through inanimate pathways: a New Zealand per-
mate change-induced productivity changes in European forests? Environ.
spective. Rev. Sci. Tech. 29, 123–133.
Res. Lett. 12, 034027.
Turvey, N.D., Attiwill, P.M., Cameron, J.N. and Smethurst, P.J. 1983
Richardson, B., Kimberley, M., Ray, J.W. and Coker, G.W. 1999 Indices of
Growth of planted pine trees in response to variation in the densities of
interspecific plant competition for Pinus radiata in the Central North
naturally regenerated acacias. For. Ecol. Manage. 7, 103–117.
Island of New Zealand. Can. J. For. Res. 29, 898–905.
Walther, G.-R., Roques, A., Hulme, P.E., Sykes, M.T., Pyšek, P., Kühn, I.,
Ridley, G., Bain, J., Bulman, L., Dick, M. and Kay, M. 2000 Threats to New
et al. 2009 Alien species in a warmer world: risks and opportunities.
Zealand’s indigenous forests from exotic pathogens and pests. Sci.
Trends Ecol. Evol. 24, 686–693.
Conserv. 142, 143.
Watson, M., Kriticos, D., Drayton, G., Teulon, D. and Brockerhoff, E. 2008
Robinet, C., Baier, P., Pennerstorfer, J., Schopf, A. and Roques, A. 2007
Assessing the effect of Essigella californica on Pinus radiata at two sites
Modelling the effects of climate change on the potential feeding activity
in New Zealand. N. Z. Plant Prot. 61, 179–184.
of Thaumetopoea pityocampa (Den. & Schiff.) (Lep., Notodontidae) in
Watt, M.S., 2003. Modelling the Influence of Weed Competition on
France. Glob. Ecol. Biogeogr. 16, 460–471.
Growth of Juvenile Pinus radiata at a dryland site. In, Forestry. University
Rogelj, J., Meinshausen, M. and Knutti, R. 2012 Global warming under
of Canterbury, Christchurch, New Zealand.
old and new scenarios using IPCC climate sensitivity range estimates.
Watt, M.S., Kirschbaum, M.U.F., Paul, T.S.H., Tait, A., Pearce, H.G.,
Nat. Clim. Change 2, 248.
Brockerhoff, E.G., et al., 2008. The effect of climate change on New
14
Climate change effects on plantation forests
Zealand’s planted forests: Impacts, risks and opportunities. Client Report
Wolken, J.M., Hollingsworth, T.N., Rupp, T.S., Chapin, F.S., Trainor, S.F.,
No. CC MAF POL_2008-07 (106-1) -No. 1.
Barrett, T.M., et al. 2011 Evidence and implications of recent and pro-
Watt, M.S., Kriticos, D.J. and Manning, L.K. 2009 The current and future
jected climate change in Alaska’s forest ecosystems. Ecosphere 2,
potential distribution of Melaleuca quinquenervia. Weed Res. 49, 381–390.
1–35.
Downloaded from https://academic.oup.com/forestry/article/92/1/1/5065106 by New Zealand Forest Research Institute Ltd. user on 09 December 2020
Watt, M., Kriticos, D., Potter, K., Manning, L., Tallent-Halsell, N. and
Woodall, S. 1983 Establishment of Melaleuca quinquenervia seedlings
Bourdot, G. 2010 Using species niche models to inform strategic man-
in the pine-cypress ecotone of southwest Florida. Florida Sci. 46,
agement of weeds in a changing climate. Biol. Invasions 12, 3711–3725.
65–72.
Watt, M.S., Palmer, D.J. and Bulman, L.S. 2011a Predicting the severity of
Wratt, D.S., Tait, A., Griffiths, G., Espie, P., Jessen, M., Keys, J., et al. 2006
Dothistroma needle blight on Pinus radiata under future climate in New
Climate for crops: integrating climate data with information about soils
Zealand. New Zealand. J. Forestry Sci. 41, 207–215.
and crop requirements to reduce risks in agricultural decision-making.
Meteorol. Appl. 13, 305–315.
Watt, M.S., Palmer, D.J. and Bulman, L.S. 2011b Predicting the severity of
Dothistroma on Pinus radiata under current climate in New Zealand. For.
Ziska, L.H. 2011 Global climate change and carbon dioxide: assessing
Ecol. Manage. 261, 1792–1798.
weed biology and management. In Handbook on Climate Change and
Agroecosystems: Impacts, Adaptation, and Mitigation. H. Daniel and R.
Watt, M.S., Palmer, D.J., Bulman, L.S. and Harrison, D. 2012a Predicting
Cynthia (eds). Imperial College Press, pp. 191–208.
the severity of Cyclanuesma needle cast on Pinus radiata under future
climate in New Zealand. J. Forestry Sci. 42, 65–71.
Ziska, L.H., Blumenthal, D.M., Runion, G.B., Hunt, E.R. and Diaz-Soltero, H.
2011 Invasive species and climate change: an agronomic perspective.
Watt, M.S., Rolando, C.A., Palmer, D.J. and Bulman, L. 2012b Predicting
Clim. Change 105, 13–42.
the severity of Cyclaneusma minus on Pinus radiata under current cli-
mate in New Zealand. Can. J. For. Res. 42, 667–674.
Ziska, L.H. and McConnell, L.L. 2015 Climate change, carbon dioxide,
and pest biology: monitor, mitigate, manage. J. Agric. Food Chem. 64,
Withers, T. and Keena, M., 2001. Lymantria monacha (nun moth) and L.
6–12.
dispar (gypsy moth) Survival and Development on Improved Pinus radiata.
15
Document Outline