


 The effects of tahr in
alpine and subalpine
ecosystems
A summary of potential and alternative monitoring networks to assess the
ecological integrity of subalpine and alpine vegetation exposed to tahr grazing
The effects of tahr in
alpine and subalpine
ecosystems
A summary of potential and alternative monitoring networks to assess the
ecological integrity of subalpine and alpine vegetation exposed to tahr grazing



 What we know about
Infrastructure available to report on the effects
the effects of tahr in
of tahr on alpine and subalpine ecosystems:
alpine and subalpine
What we know about
Infrastructure available to report on the effects
the effects of tahr in
of tahr on alpine and subalpine ecosystems:
alpine and subalpine
› Plots in alpine grasslands (n = 117) in 8 catchments in the management zone,
ecosystems:
established in the 1990s, remeasured several times, most recently 2013;
remeasurement begins 2020.
› Alpine and subalpine
ecosystems in New Zealand
› Plots in alpine grasslands are environmentally and compositionally biased, adequate
are naïve to mammalian
for reporting change in dominant tussocks but not other less dominant species.
herbivores, so it’s likely that
› Plots in alpine grasslands are only in 8 catchments in the management zone and
these ecosystems will not
cannot be used to make inference about the whole management zone.
be resilient to some of their
effects [Forsyth et al. 2010]
› Systematically-located plots on an 8-km grid across alpine and subalpine
ecosystems across the management and exclusion zones, established 2011–2017;
› Tahr are social animals that
remeasurement began in 2018.
affect ecosystems differently
› Systematically-located plots are adequate to report on change in structural
in space and time: alpine
dominant species (tussocks, shrubs) but not for other less dominant species (e.g.,
ecosystems in summer
small herbs eaten by tahr).
and subalpine ecosystems
(especially shrublands) in
› Legacy datasets of transects in some alpine and subalpine ecosystems on public
winter [Forsyth & Tustin 2005]
conservation land (established 1970s/80s), not remeasured since.
› Legacy datasets of transects in subalpine grasslands on public leasehold land,
› Tahr diet in alpine and
established in 1960s/70s, measured most recently 2000s [Day & Buckley 2013].
subalpine ecosystems
consists mostly of tussocks
and shrubs but also alpine
herbs (buttercups and others)
that are naturally uncommon
and patchy in distribution
[Tustin & Parkes 1988; Parkes
& Forsyth 2008]
› Tahr can have highly
concentrated effects in alpine
and subalpine ecosystems,
transforming tall tussocks
and subalpine shrublands to
turfs at local (<300 m2) scales
[Wilson 1976; Wardle 1977,
1979]
› As tahr abundance increases,
the heights of dominant
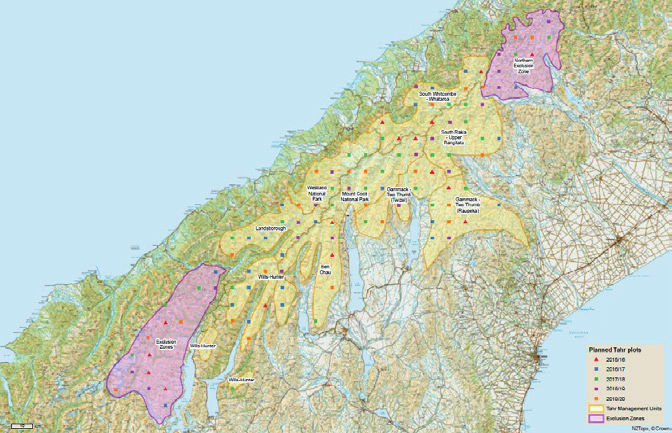
▲ Tier 1 20 x 20 plots within the tahr management units and exclusion zones as defined in the Tahr
tussocks decline at wider
Control Plan.
scales [Cruz et al 2017]
▲ Vegetation in one of the 117 subjectively located plots (in Zora Creek, Westland) at two points in
time showing a reduction in tussock cover and height. Left image in 1999; right image in 2012.
3
Photos: Ingrid Grüner, DOC
Cover image: Juvenile males.
Photo: Dylan Higgison



 Here’s the state of our capacity to report on the effects of tahr:
Here’s the state of our capacity to report on the effects of tahr:
› We can report on change in alpine tussock grasslands
from plots established between 1990 and 1999. These
(d)
plots showed a small but significant decline in tussock
grass height over time [Cruz et al 2017]
› New systematically-located plots (established since
2011) provide a baseline against which changes in alpine
and subalpine ecological integrity can be determined.
These plots have established that there is lower shrub
cover in the tahr management zone than the exclusion
zone [Bellingham et al 2018]
› So far, reporting of effects of tahr on ecosystems is from
public conservation land only.
1990
1995
2000
2005
2010
Year
▲ Image of figure from
Cruz et al showing change in tussock height.
We cannot report on:
› Resilience of alpine and subalpine species and
ecosystems to grazing by tahr.
› Whether tahr are impoverishing vegetation widely or
transforming ecosystems widely because we lack time-
series data.
› Effects on herbaceous species eaten by tahr. We need
more information to inform what groups of plants to
monitor and why. Our capacity to select other species to
measure is currently statistically poor and hampered by
a lack of knowledge about tahr’s dietary preference (we
know that herbaceous plants like Ranunuculus species
comprise up to a quarter of their diet by dry weight
[Parkes & Forsyth 2008], but we are unable to determine
whether this affects their populations).
› Catchment-scale impacts of tahr and ecosystem
resilience that integrate tahr behaviour and movements
with vegetation changes in alpine and subalpine
ecosystems.
▲ Herbaceous species.

 References
Things that could better inform
References
Things that could better inform
Bellingham PJ, Wiser SK, Burge OR, Easdale T, Richardson SJ 2018.
decisions about the ecological impacts
Potential of Tier One and alternative monitoring networks to
assess the ecological integrity of alpine vegetation exposed to
of tahr and how to set thresholds for
tahr grazing. Landcare Research Contract Report LC3328 for the
intervention that are scientifically
Department of Conservation.
defensible for both alpine and
Cruz J, Thomson C, Parkes JP, Grüner I, Forsyth DM 2017. Long-
subalpine ecosystems:
term impacts of an introduced ungulate in native grasslands:
Himalayan tahr (Hemitragus jemlahicus) in New Zealand’s
› Continue measurement of existing plots in alpine
Southern Alps. Biological Invasions 19: 339–349.
tussock grasslands in the management zone
Day NJ, Buckley HL 2013. Twenty-five years of plant community
and of systematically-located plots across the
dynamics and invasion in New Zealand tussock grasslands.
management and exclusion zones.
Austral Ecology 38: 688–699.
› Establish new networks of plots (or reinstate
Forsyth DM, Tustin KG 2005. Himalayan tahr. In: King CM (ed) The
old plot networks) in alpine and subalpine
Handbook of New Zealand Mammals, 2nd edition, pp. 361–373.
ecosystems to increase statistical power to
Oxford University Press, South Melbourne.
detect tahr browsing effects.
Forsyth DM, Wilmshurst JM, Allen RB, Coomes DA 2010. Impacts of
introduced deer and extinct moa on New Zealand ecosystems.
› Instigate long-term research sites that collect
coupled data on tahr and other mammalian
New Zealand Journal of Ecology 34: 48–65
herbivores with vegetation data in alpine
Parkes JP, Forsyth DM 2008. Interspecific and seasonal dietary
grasslands and subalpine grasslands that are
differences of Himalayan tahr, chamois and brushtail possums
heavily browsed by tahr east and west of the
in the central Southern Alps, New Zealand. New Zealand Journal
Divide.
of Ecology 32: 46–56.
Tustin KG, Parkes JP 1988. Daily movement and activity of female
› Maintain current methods used on plots to
and juvenile Himalayan tahr (Hemitragus jemlahicus) in the
determine effects of tahr (to ensure continuity
eastern Southern Alps, New Zealand. New Zealand Journal of
and ability to maximise time-series data)
Ecology 11: 51–59.
and evaluate suitability of other methods to
determine effects.
Wardle P 1977. Plant communities of Westland National Park (New
Zealand) and neighbouring lowland and coastal areas. New
› Determine the effectiveness of multiple remote
Zealand Journal of Botany 15: 323–398.
sensing techniques to determine tahr habitats
Wardle P 1979. Plants and Landscape in Westland National Park.
and their impacts on them.
National Parks Authority Scientific Series No. 3, Department of
› Determine dietary preference of tahr (i.e.,
Lands and Survey, Wellington.
ascertain biomass of plant species in the wild
Wilson HD 1976. Vegetation of Mount Cook National Park, New
in proportion to their mass in tahr rumens),
Zealand. National Parks Authority Scientific Series No. 1,
including throughout seasons.
Department of Lands and Survey, Wellington.
› Improve the evidence base for attributing impacts
on vegetation to tahr, in the context of other
herbivores. This would include determining hare
diets in alpine or subalpine ecosystems .
› Determine targets (optimum ecological integrity)
to which to management of alpine and subalpine
ecosystems could be aimed.
Photo: Scott Theobald (DOC)
1